A kérészek (Ephemeroptera) rendje viszonylag kicsi, kb. 3330 ismert fajukat 440 nemzetségbe és 40 családba sorolják. Világszerte körülbelül 1000 faj vár még leírásra. Az ismert fajok többsége a Baetidae (956), a Heptageniidae (598) és a Leptophlebiidae (643) családba tartozik. Már a karbon korban is létezett rovarcsoport egykor sokkal fajgazdagabb volt. Sok kérészfaj maradványait megtalálták borostyánban. Egyes feltételezések szerint ennek az lehet a legfőbb oka, hogy a gyantától ragacsos kidölt fatörzsek vízszintesen poláros fényt tükröztek, ami megtévesztette és magához vonzotta a pozitív polarotaxissal vizet kereső vízirovarokat, mint például a kérészeket.
A törékeny testű áttetsző szárnyú rovarok nem táplálkoznak, többnyire csak néhány napig élnek. Kifejlett alakjuk rövid életére utal tudományos nevük is (Ephemeros: egy napig élő, pteron: szárny). Szűkre szabott életük egyetlen feladata az utódnemzedék létrehozása. A kifejlett állatok fején rövid csápok és nagy összetett szemek vannak (1-2. ábrák). A Baetidae és Leptophlebiidae család hím egyedeinél turbánszem alakul ki, amely egy terjedelmes turbánalakú felső és egy alatta elhelyezkedő gömbölyű összetett szemből áll. A turbánszem felső részével ismerik fel a hímek a nőstényeket az alkonyati égbolt világos hátterénél, alulról megközelítik őket, majd a levegőben párzanak velük. A kérészek hártyás szárnyaikat nyugalmi helyzetben összecsapják a testük fölött. Az alsó permből származó első kérészeknek még két pár egyforma szárnyuk volt, de a harmadidőszaki Balti borostyánkőből kimutatott fajoknál már a hátsó szárny redukcióját figyelhetjük meg. A 2. pár szárny néhány fajnál (Cloeon, Caenis) teljesen elcsökevényesedett. A tor három szelvényből áll, ezek viselik a lábakat. A hímeknél az első lábpár rendszerint hosszabb, a középső és a hátsó lábpár viszont egyforma hosszúságú. Lábak közül az elsők működésükben helyettesíthetik az igen rövid csápokat. A potroh 10 szelvényből áll. A hímeknél a 9. szelvény ventrális oldalán a több ízből álló ivarlábak és a középső dorzális részen pedig a penis nyúlványok tűnnek a szemünkbe. A hím ivarkészülékek alakja fajonként változó, jó határozóbélyeg. A nőstények ivarnyílása a 7-8. potrohszelvény ventrális oldalán nyílik.

1. ábra Kérész (Ephemeroptera) szubimágók. (a) Erezett kérész (Heptageniidae) – 12 mm. (b) Hím Baetis rhodani szubimágó – 1 cm
A testvégen két vagy három hosszú, fonálszerű nyúlvány található. A két szélső a farktoldalék, míg a néha hiányzó középső, a végfonál.
A kérészek testhossza 3-35 mm között változik. A kifejlett egyedek nem táplálkoznak, bélcsatornájukat levegővel pumpálják fel, ezért potrohuk gyakran üvegszerűen áttetsző. A fajok többsége az esti órákban rajzik. Ilyenkor az utolsó lárvaalak, a nimfa a vízfelszínre úszik, vagy kimászik a szárazra, ahol néhány perc alatt előimágóvá, szubimágóvá vedlik. Az előimágó a kifejlett kérészhez hasonló szárnyas rovar, amely még egy vedlést követően válik ivarérett imágóvá (1-3., 15. és 25. ábra).

2. ábra (a-b) Erezett- (Heptageniidae) és teleszkópszemű (Baetidae) kérész szubimágók

3. ábra Dunavirág (Ephoron virgo, Polymitarcyidae) szubimágó imágóvá való vedlése
A szubimágók színei kevésbé élénkek szárnyaik opálosak, farktoldalékaik rövidebbek, mint az imágóké. A szubimágók nem minden faj esetében jelennek meg, a tiszavirágnál (Palingenia longicauda) például csak a hímeknek van szubimágó alakja, a lárvabőrből kibújó nőstények rögtön párzóképesek. A kérészeket rajzásuk helye alapján három csoportba sorolhatjuk: mindvégig a víz fölött rajzó fajok; szárazföld fölött rajzó, de a vízzel vizuális kontaktusban maradó fajok; a víztől távol rajzó, a vízfelszínnel vizuális kapcsolatot nem tartó fajok. A szárazföld fölötti kérészrajzáskor a hímek általában kisebb csapatokba verődnek egy jól látható tereptárgy körül és megkezdik felfelé emelkedő, majd süllyedő násztáncukat.
A rajokba repülő nőstényekre a hímek alulról csapnak le és párzanak velük. A megtermékenyítést követően a nőstények kitojják petecsomójukat. Egy petecsomó gyakran 6-8000 petét is tartalmaz. A petés nőstények a vízhez repülnek, hogy a potrohvégükön csüngő petecsomójukat a vízbe juttassák. Egyes fajoknál a nőstények jelentős, akár 50%-os aránya is szűznemzéssel szaporodik. A megtermékenyítetlen petékből csak nőstények fejlődnek ki. Néhány kérészfajnál az elevenszülés (ovoviviparia) is előfordul (pl. Cloeon dipterum), amikor a nőstények nem petéket, hanem lárvákat juttatnak a vízbe. Az áramló vizekben élő fajoknál a nőstények a folyásiránnyal ellentétesen repülnek a víz fölött, mielőtt a vízfelszínre ereszkedve lerakják petéiket. E kompenzációs repülésükkel a vízbe kerülő peték elsodródását ellensúlyozzák.
A kérészek a vizet polarotaxis segítségével találják meg, amely a vízfelszínről visszavert, vízszintesen polarizált fény érzékelésén alapszik. Nemrégiben sikerült kimutatni, hogy a dunavirág (Ephoron virgo) és egy törpekérész faj a Caenis robusta számára a vízszintesen polarizált fény sokkal vonzóbb, mint a polarizálatlan fény, és a függőlegesen polarizált fény a legkevésbé vonzó, ha az inger intenzitása és spektruma azonos. Ez a kérész viselkedés biztosíthatja a rajzás kialakulását a vízfelszín felett.
A vízszintesen polarizált fényt tükröző nyílt vízfelület mellett a partközeli vízfelszínre vetülő növényzeti árnyék és tükörkép gyengén és nem vízszintesen (főleg függőlegesen) polarizált fényt tükröz. Emiatt a vízfelszín nyílt részei felett rajzó polarotaktikus kérészek folyamatosan itt tartózkodnak, és elkerülik a víztest polarizálatlan vagy nem vízszintesen poláros fényt tükröző peremterületeit.
A rovarok gyakran rajzanak aszfaltutak, sötét színárnyalatú gépkocsik és fényes-fekete agrofólia fölött, amelyekre sokszor a petéiket is lerakják a nőstények (4-5. ábrák). Ez a napnyugta körül kialakuló természetellenes viselkedés azzal magyarázható, hogy a felsorolt mesterséges objektumok felületéről visszaverődő fény polarizációs iránya ebben az időszakban hozzávetőlegesen vízszintes, polarizációfoka pedig gyakran magasabb, mint a vízfelszínről visszaverődő fényé. Ennek következtében a mesterséges felületek a vízfelületnél vonzóbbak lehetnek a vizet kereső polarotaktikus kérészek számára.
A patakokból kirepülő dánkérész (Ephemera danica) rajzása során kimutatták, hogy a nőstények folyásiránnyal szembeni kompenzációs repülése eltérülhet olyan hidaknál, melyeken aszfaltút fut. Az aszfaltút viszonylag gyenge, vízszintesen polarizált jelzése önmagában nem vált ki petézést, de hatására megjelenik a kérészek vízkövető viselkedése. Így a patakon átvezető híd aszfaltútja képes a dánkérész (E. danica) nőstényeket egy távoli, erősen és vízszintesen poláros mesterséges felülethez vezetni, amelyek petézést váltanak ki. Ezért a polárosan fényszennyező aszfaltút elvezetheti a petézés előtt álló kérészeket a patakokból egy erősen és vízszintesen poláros felülethez, például egy parkoló fényes, sötét színű autóhoz, ami már képes kiváltani a rovarok petézését. Hasonló jelenség figyelhető meg a Duna egyik mellékfolyója, az Ipoly mentén. Itt a folyón átívelő hídtól, az út felett 3-5 m-es magasságban repülve hagyják maguk mögött a folyót a nyár elején rajzó tarka kérész (Ephemera vulgata) egyedek. Ilyenkor a kérészek az aszfaltutat követve 3-4 kilométerre is eltávolodhatnak az Ipolytól, ahol le kellett volna rakni a tojáscsomóikat.

4. ábra (a) Nőstény tiszavirág (Palingenia longicauda) száraz aszfaltúton. (b) Tiszavirág lerakott tojásaival nedves aszfaltúton

5. ábra Kérészek fénypolarizáló autókarosszérián. (a) Egy nőstény és három hím Baetidae imágó vörös autókarosszérián – 9 mm. (b) Hím teleszkópszemű kérész (Baetis rhodani, Baetidae) imágó autószélvédőn – 7 mm. (c) Vörös autókarosszériára petéző dánkérész (Ephemera danica, Ephemeridae) – 20 mm
A poláros fényt tükröző felületek a folyó fölött rajzó kérészekre is veszélyt jelenthetnek, mert például a sötétedés után rajzó dunavirág (Ephoron virgo) több millió egyedét vonzhatják magukhoz a hidak lámpái, ahol a nőstények a víz helyett a híd aszfaltútjára rakják le petéiket. A kivilágított hidak kettős optikai hatáson alapuló ökológiai csapdát képeznek. Az erős fényű hídlámpáik pozitív fototaxissal nagyobb távolságból is magukhoz vonzzák a folyó fölött a kompenzációs repülésüket folytató nőstény kérészeket (6. ábra). A hídon pedig az aszfaltút poláros jele pedig leszállásra és petézésre készteti a rovarokat (7. ábra). A hídon elpusztuló óriási rovartömeg sem a halaknak sem a vízparti békáknak nem szolgál táplálékul, legfeljebb a denevérek és a hídon élő pókok hasznosítják ezeket táplálkozásuk során (8. ábra).

6. ábra (a-c) Milliónyi dunavirág (Ephoron virgo) nőstény a tahitótfalui Duna-hídon

7. ábra Dunavirág (Ephoron virgo) nőstények és sárga petecsomóik az aszfaltúton
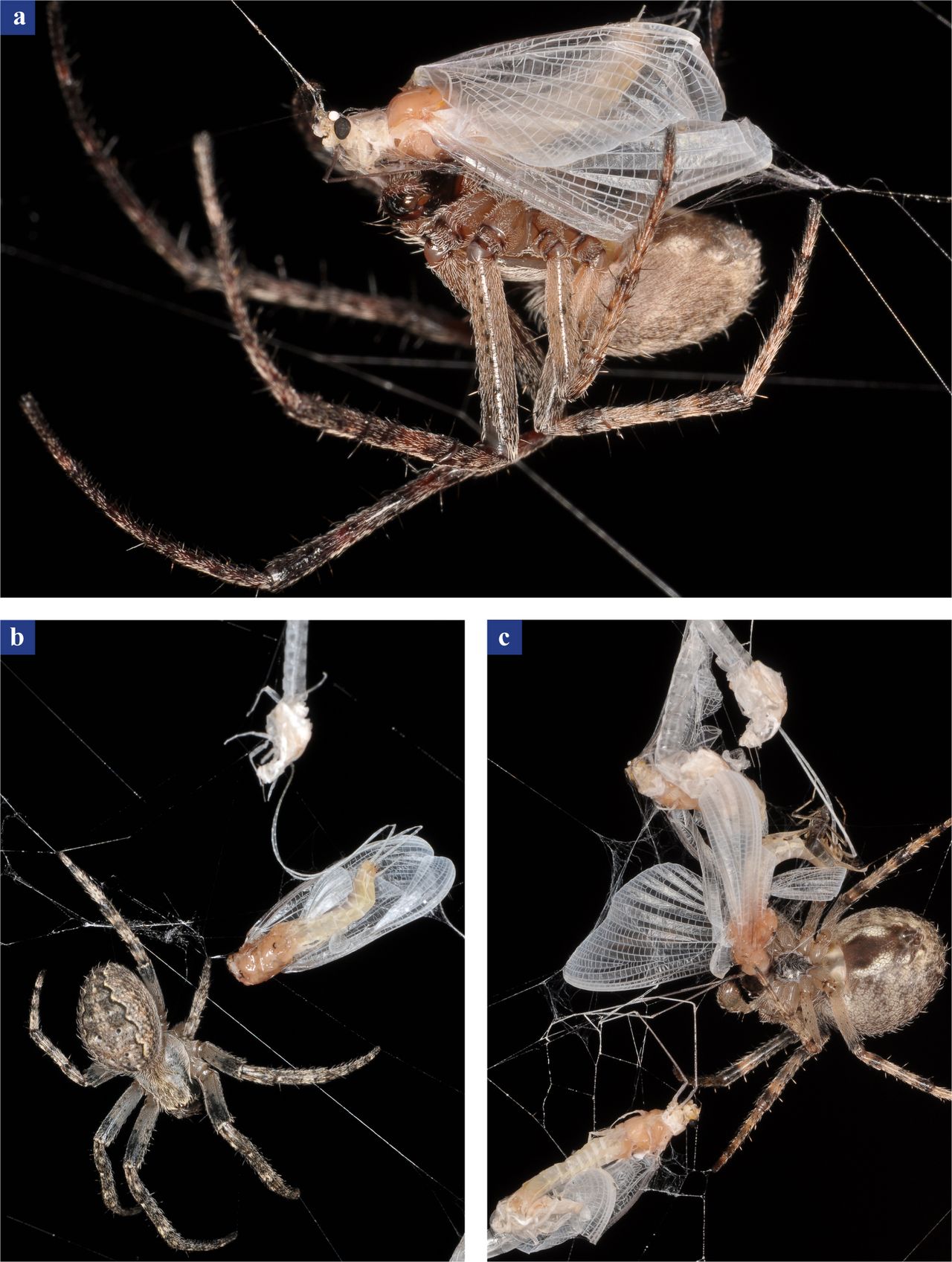
8. ábra Keresztespók dunavirág szubimágókat és nőstány imágókat zsákmányol
A hidak természetkárosító hatásának kivédésére sikeresen tesztelésre került egy módszer, ami a kérészek pozitív fototaxisát kihasználva védi az utódgenerációt.
Terepkísérletek során hídról lelógatott, a folyás irányába világító reflektorokkal sikerült a víz fölött tartani, majd a vízbe csalogatni a peterakó nőstényeket. Így meg lehetett akadályozni, hogy a hídon lévő, vagy a parti lámpákhoz vonzódjanak, és a szárazon pusztuljanak el petecsomóikkal együtt. A kérészek folyótól való elrepülését, az idő és a lelógatott reflektorok ki és bekapcsolt állapotának függvényében mérve megállapítást nyert, hogy ez a módszer gyakorlatilag teljes mértékben képes megakadályozni a kérészek pusztulását. Így különös módon a dunavirág példáján sikerült demonstrálni, hogy az éjjel rajzó kérészek utódgenerációja miként védhető meg a petecsomók pusztulását okozó mesterséges fényektől fény segítségével (9. ábra).

9. ábra Kérészvédő fénysorompó a rábahídvégi Rába-hídnál. (a) A fénysorompó lámpái a hídhoz érkező kérészrajt világítják meg. (b-c) A rábahídvégi híd a ki- és bekapcsolt fénysorompóval. (d) A fény feltartóztatja a kérésztömeget, amely a vízbe kerülve elsodródik
A világon elsőként 2019 tavaszán a tahitótfalui Tildy Zoltán hídon épült ki egy spektrálisan optimalizált kérészvédő fénysorompó, amely képes megakadályozni a védett dunavirág tömeges pusztulását a szárazföldön. A kérészvédő fénysorompó alkonyatérzékelő relé kapcsolóval és időzítővel van felszerelve, ami lehetővé teszi az eszköz automatikus működtetését a dunavirág rajzásóráira korlátozva (10. ábra).

10. ábra (a) Kérészvédő fénysorompó felszerelése a tahitótfalui Tildy Zoltán hídra 2019 tavaszán. (b) A kékfényű fénysorompó müködés közben
A kérészek petéiből kikelő lárva 05,-3 éves fejlődése során 15-16-szor vedlik és a szárnyhüvelyek nagysága, színezete alapján 5 fejlődési stádiumot különíthetünk el. A lárvák Közép-Európában elsősorban az áramló vizek lakói, csak a Cloeon és a Caenis fajok élnek állóvizekben is.
A lárvatest elején a fejet találjuk, amelyen felismerhetők az összetett szemek, a két csáp és a rágótípusú szájszervek. A fej mögött a háromszelvényű tor következik, amely a lábakat viseli. Az idősebb lárváknál a középső szelvény dorzális részén a szárnyhüvelyek figyelhetők meg, amelyekben a szárnyak fejlődnek. A fejlett nimfák (utolsó stádiumú lárvák) elülső szárnyhüvelyei sötét színűvé válnak az átváltozás közeledtével.
A tor ventrális oldalán találhatjuk a járóláb típusú ízelt lábakat. A potroh 10 szelvényből áll és a lemez, vagy pamacs alakú tracheakopoltyúkat viseli. A kopoltyúpárok száma rendszerint 7 pár és alakjuk az egyes családokra, fajokra jellemző. A potrohon lévő kopoltyúk mellett ritkán az alsó álkapocs tövében is lehetnek ilyen légzőszervek (Oligoneuiidae). Az utolsó szelvényen 2 vagy 3 hosszú potrohfüggeléket találunk.

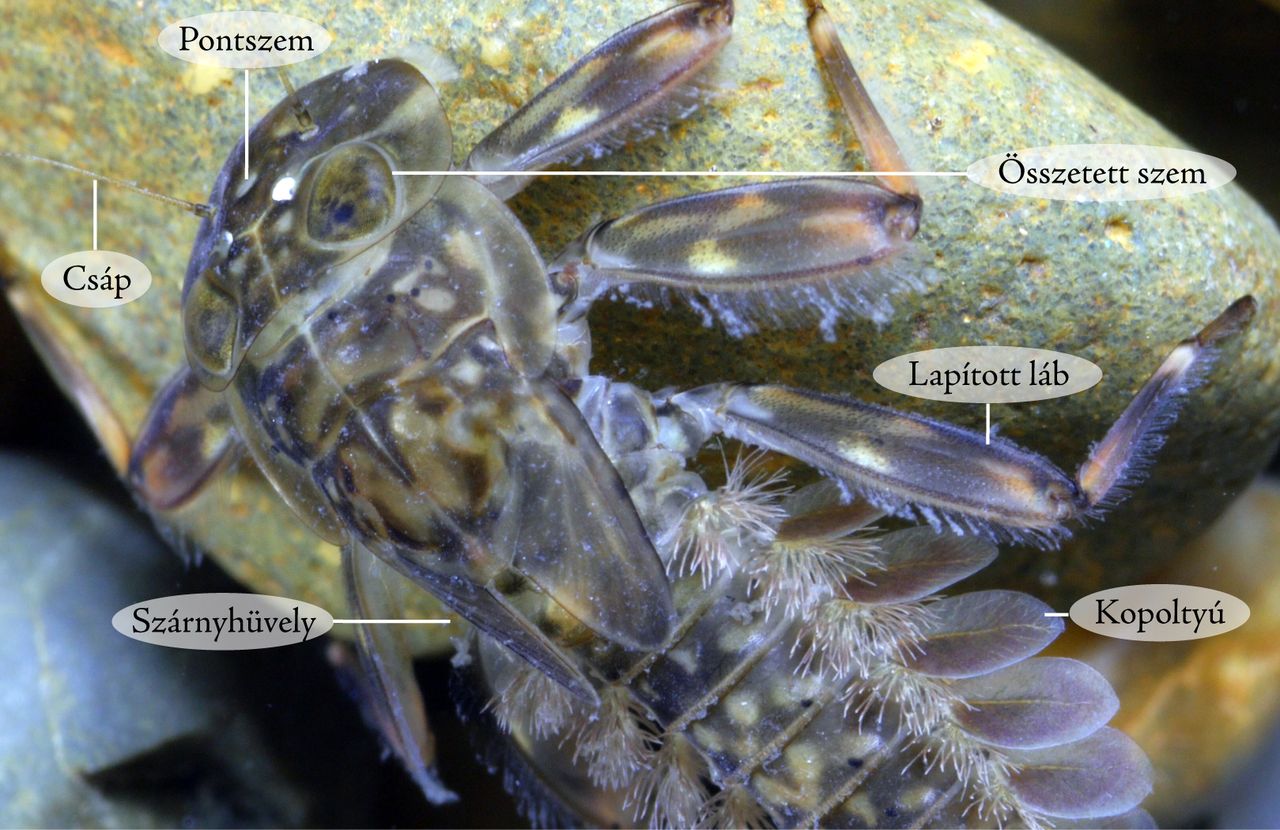
11. ábra Epeorus assimilis (Heptageniidae) lárva testfelépítése – 1 cm
12. ábra Lapított testű kérészlárva (Heptageniidae)
A lárvák különböző vízsebességekhez alkalmazkodva eltérő testalakot vehetnek fel. A dorzoventrálisan lapított, fedetlen kopoltyús fajok (pl. Epeorus, Ecdyonurus, Rhithrogena) rohanó hegyipatakokban élnek (11-12. ábrák). A hengeres testalkatú fajok, mérsékeltebb vízfolyásnál vagy állóvizekben élő állatok (pl. Cloeon, Baetis, Potamanthus). A lapítottabb, kopoltyúfedős fajok, lassúfolyású, vagy állóvízi körülmények között, az iszapfelszínen és az iszapban élnek (pl. Ephemerella, Caenis).
A patakok és folyók üledékében élnek az ásó életmódot folytató lárvák (Palingenia, Ephemera, Ephoron) (13., 16. és 26. ábrák). Erős lábaik, szorosan a testhez simulnak. Fonalas tracheakopoltyúik ferdén hátra hajolva a hátoldalon helyezkednek el, hogy ne akadályozzák az iszapban való előrehaladást. Elülső lábaik ásásra módosultak és az erős rágók szintén hasznos ásó-, és fúrókészülékek. Táplálékként a hordalék szerves anyagát, apró élőlényeket, baktériumokat, mikroalgákat hasznosítanak. A kérészlárvák biomasszája a folyóvizekben, hegyi patakokban meghaladhatja az árvaszúnyog lárvákét, ezért igen fontos haltáplálék szervezetek.

13. ábra Dunavirág (Ephoron virgo, Polymitarcyidae) lárvák ásás közben – 15 mm
A kérészlárvákon és testükben különböző bakteriális, protoza, lapos- és fonálféreg élősködők élnek. A kérészlárvák halparaziták köztes gazdái vagy az élősködők végső gazdái lehetnek. Egyes árvaszúnyog (pl. Symbiocladius rhithrogenae) lárvák is lehetnek kérészlárvák ektoparazitái, miközben a gazda hemolimfájával táplálkoznak, és sterilitást okozhatnak (14. ábra).

14. ábra (a-b) Élősködő árvaszúnyog (Symbiocladius rhithrogenae) lárva (a) és báb (b) erezett kérészlárva (Electrogena lateralis, Heptageniidae) szárnyhüvelyében – 4 mm
A Prosopistomatidae család egyetlen európai faja a Prosopistoma pennigerum. A folyókban élő lárva 8 mm-es, pajzs alakú.
Gyorsfolyású patakokban és hegyi folyókban élnek az erezett kérészek (Heptageniidae) lárvái (11-12. és 24. ábrák). A lárvák a vízben heverő kőgörgetegek algabevonatát legelik, miközben teljesen a felszínéhez lapulnak. Élőhelyükön fokozottan jelentkezik az elsodródás veszélye, ami ellen erőteljesen lapított testalkatukkal védekeznek. Potrohuk két oldalán levélalakú tracheakopoltyúk találhatók, amelyek tövében vékony szálacskák növelhetik tovább a légzőfelületet.
Az Arthropleidae család egyetlen ritka európai faja az Arthroplea congener. Szerves törmelékkel táplálkozó lárvája a folyók, patakok és átöblítődő kis tavak partközeli részén a vízinövényzetbe, kövekbe kapaszkodva él. Az egyéves fejlődésű faj májusban, júniusban rajzik, a telet tojás formában vészeli át. A lapított testű lárva széles fejének hátoldalán méretes összetett szemek vannak. Nagy kefeszerű, a fejen túlnyúló állkapcsi tapogatóik (palpus maxillaris) alapján jól megkülönböztethetők minden más kérészlárvától.
A tiszavirág–félék (Palingeniidae) lárvái a fenék üledékében élő sárgásfehér színű, hengeres testű állatok. Erőteljes rágóikkal és ásólábaikkal fúrnak járatokat az aljzatban. Legismertebb fajuk a Tiszában a Rábában, a Dnyeszterben, Prutban és mellékfolyóikban élő tiszavirág (Palingenia longicauda) (15. és 26b. ábrák). A lárvák három évig fejlődnek a meder agyagpadjaiban, majd egy kora nyári alkonyon hat óra körül a nimfák a vízfelszínre úsznak, és szárnyas rovarrá vedlenek. A hím egyedek szubimágók, amelyek a partra repülnek, még egyszer vedlenek és így alakulnak ivarérett imágóvá (15. ábra). A nőstények a lárvabőrből kibújva rögtön alkalmasak a párzásra. A nőstények a rajzás során mindvégig a vízfelszín fölött maradnak. A nászrepülés és a párzás is a víz fölött történik, ami után a nőstények megkezdik kompenzációs repülésüket. Ennek során néhány kilométert repülnek a folyásiránnyal szemben, majd a vízfelszínre ereszkedve lerakják petéiket. A vízárammal lefelé sodródó peték elérik a szülők bölcsőjét jelentő agyagpadot biztosítva három év múlva az újabb generáció kirepülését. A mintegy kétórás rajzás után a 4 cm-es rovarok milliónyi sárgás teteme virágszőnyegként borítja a folyó habjait. A nagytestű rovarok kedvenc csemegéi a madaraknak és halaknak, ezért a faj biztos fennmaradását csak a tömegrajzás kialakulása biztosíthatja. Ezért is van fokozott veszélynek kitéve a tiszavirág a folyószennyezéseknek és a rajzásidőben kialakuló magas vízállásoknak.

15. ábra Tiszavirág (Palingenia longicauda, Palingeniidae) átváltozása. (a) Nőstény imágó kibújása a lárvabőrből. Az állat a vízfelszínről lett kiemelve. (b-d) Hím szubimágó vedlése imágóvá egy vízparti csónakon – 40 mm
A tarka kérészek (Ephemeridae) lárvái az áramló vizek aljzatában élő 30 mm-es krémszínű állatok. Ásásra módosult ízelt lábaik erőteljesek, rágóik jellegzetesen kifelé hajlók. Tracheakopoltyúik a potroh hátoldalára hajlanak. Gyakori fajaik a dán kérész (Ephemera danica) (16a-b. és 26a. ábrák) és a tarka kérész (E. vulgata).
A dunavirág–félék (Polymitarcyidae) családjába sorolt dunavirág (Ephoron virgo) a tiszavirághoz hasonló életmódot folytat és sokszor vele egy élőhelyen fordul elő. A folyók üledékében élő lárva hasonlít a tarkakérészek lárváihoz, de azoknál kisebb, legfeljebb 15 mm-es és rágóinak hegye nem kifelé, hanem a test középvonala felé hajlik (13. és 26c. ábrák). Az imágók július végén és augusztusban rajzanak. A lerakott peték tavasszal indulnak fejlődésnek, a lárvák április közepén kelnek ki. Az Ephoron virgo korábban évtizedekre eltűnt a legtöbb szennyezett közép-európai folyóból, például a Rajnából és a Dunából. A vízminőség javulásának köszönhetően a 1990-es években jelentek meg újra tömegrajzásaik.

16. ábra (a) Dán kérész (Ephemera danica, Ephemeridae) ásó típusú lárvája – 3 cm. (b) Fekete fóliára petéző dán kérész (kis kép) és lárva a vízfenéken. (c) Dunavirág (Ephoron virgo, Polymitarcyidae) lárva – 15 mm
A folyamvirág–félék (Potamanthidae) családjának egyetlen képviselője él Európában a folyamvirág (Potamanthus luteus). A szárnyas rovarok sárgák. Az áramvonalas testű 18 mm-es lárva rágói felülnézetből nem láthatóak és tracheakopoltyúik oldalt állnak. Ezek tollszerűek és 7 pár van belőlük. Az első pár elcsökevényesedhet. A folyamvirág lárvák kisebb és nagyobb folyók partközeli részein élnek a kövek és detritusz között (17. és 28a. ábrák).

17. ábra Folyamvirág (Potamanthus luteus, Potamanthidae) lárva – 17 mm
A törpekérészek (Caenidae) családjába zömök testű rovarok tartoznak. A lárvák aprók, testhosszuk nem haladja meg a 9 mm-t. A tracheakopoltyúik közül az első pár apró, alig észrevehető. Ezzel szemben a második pár igen nagy, elfedi az összes további légzőszervet. A törpekérész lárvák testét szinte mindig iszapszemcsék, és finom törmelék borítja (18. és 27b. ábrák). Az elevenszülő kérészhez hasonlóan egyes fajaik állóvízben élnek és a kifejlett rovaroknak csak egy pár szárnya van.

18. ábra Törpekérész (Caenidae) lárvák – 9 mm
A Neoephemeridae családnak egyetlen európai faja a Neoephemera maxima. A tiszta, áramló vizet kedvelő lárvák többnyire a víz alatti fatuskókon kapaszkodnak. A lárva 2-5 tracheakopoltyú párjainak lemezei duplák. A kopoltyúlemez belső oldalához hosszúkás rojtos lemez kapcsolódik.
Az Isonychiidae családnak egyetlen európai faja az Isonychia ignota. A lárvák felismerését az első láb tövében található kopoltyú-pamacs és a lábszár lábfej felé eső részén, a belső oldalon található 1-1 tüskeszerű nyúlvány segíti (28a-b. ábrák).
A denevérszárnyú kérészek (Oligoneuriidae) lárvái gyorsfolyású vizekben élnek. A lárvák karcsú, áramvonalas testű, széles fejű állatok. Oldalt álló kisméretű, kerek tracheakopoltyúikhoz finom szálacskák kapcsolódnak. Az első pár tracheakopoltyújuk az első potrohszelvény hasi oldalán ered.
A teleszkópszemű kérészek (Baetidae) lárvái hengeres, áramvonalas testű állatok. Potrohuk két oldalán levélszerű tracheakopoltyúk vannak. Az egyes nemzetségek elkülönítésében a fartoldalékoknak és a tracheakopoltyúknak is szerepe van. A Baetis nem fajainál a középső fartoldalék rövidebb, mint a két szélső. A tracheakopoltyúk szimplák, levél alakúak. Közép-Európában 19 fajuk él. A Cloeon lárvák levél alakú tracheakopoltyúi közül az első 6 pár páros, a 7. pedig szimpla. Közép-Európában két fajuk él álló- és lassúfolyású vizekben a Cloeon dipterum (19., 28c. és 29a. ábrák) és a C. simile. A Procloeon lárváknak szimpla, aszimmetrikus levél alakú tracheakopoltyúk vannak. Közép-Európában egyetlen fajuk a Procloeon bifidum, amely főleg növényzettel benőtt síkvidéki folyókban fordul elő. A Cloeon és Protocloeon lárvák hátulsó szárnyhüvelyei hiányoznak. A Centroptilum lárvák levél alakú tracheakopoltyúi párosak, a faroknyúlványok többé-kevésbé hasonló hosszúságúak, sötét gyűrűket viselnek. Közép-Európában 3 fajuk él. Az Acentrella génusz egyetlen közép-európai faja az A. sinaica. A középhegységi patakokban élő lárva középső testnyúlványa csökevényes 1-3 ízből áll. A két szélső nyúlvány belső oldalán nincsenek úszószőrök. A Baetopus lárvák állkapcsi tapogatója (palpus maxillaris) utolsó íze kiszélesedő és hosszú sertéket visel, a két szélső faroknyúlvány belső oldala úszószőrökkel borított. Síkvidéki folyókban élő egyetlen közép-európai faja a Baetopus tenellus, amelynek hátoldalán a 3-5. potrohszelvényen kereszthez hasonló fehér mintázat látható.

19. ábra Elevenszülő kérész (Cloeon dipterum, Baetidae ) lárvája – 6 mm
A Siphlonuridae családba tartozó kérészek lárvái robosztus megjelenésű, a 18 mm-es testhosszúságot is elérő, olajbarna színű állatok. Az európai fajok közül a tavi kérész (Siphlonurus lacustris) a leggyakoribb, amely nagyobb állóvizekben és folyókban él (20., 29b-c. és 30a-b. ábrák). Jellegzetes tulajdonságaik a potrohszelvényeikről oldalirányban kiálló tüskék, és a dupla lemezekből álló tracheakopoltyúik. A méretes levélszerű tracheakopoltyúk közül az első pár, vagy az összes duplán kapcsolódik egymáshoz. A kifejlett rovarok júniustól szeptemberig rajzanak.

20. ábra Tavi kérész (Siphlonurus lacustris, Siphlonuridae) – 1 cm
Az Ameletidae családnak két faja az Ameletus inopinatus és a Metreletus balcanicus (21. és 30c. ábrák) él Közép-Európában. Az Ameletus fajok 700 m tengerszint feletti magasságban patakokban és tavakban, míg a Metreletus fajok 500 m-nél alacsonyabban fekvő területek patakjaiban élnek. A lárva valamennyi tracheakopoltyúja egyszerű. Jellemzőjük még a bevágott alsó ajak és az állkapcson sorakozó kitintüskék.

21. ábra Metreletus balcanicus (Ameletidae) lárva – 11 mm
Az Ametropodidae család egyetlen európai faja az Ametropus fragilis. A 18 mm-es lárva első lábtövén, a belső oldalon szőrzettel borított nyúlvány található. Az elülső lábak fejletlenek, a 2. és 3. lábpárak a lábszárnál (tibia) hosszabb karomban végződnek. Az egylemezű, ovális tracheakopoltyúk peremén hosszú szőrök sorakoznak.
A Leptophlebiidae családba tartozó 12 mm-es lárváknak hosszú faroknyúlványai vannak. A 7 pár tracheakopoltyú vékony szálakban végződik. Kisebb áramló vizekben élnek az aljzat törmeléke között (22. és 31a. ábrák).

22. ábra Paraleptophlebia sp. (Leptophlebiidae) lárva – 10 mm
Az Ephemerellidae család lárvái gyors folyású patakokban és folyókban élnek. A kissé lapított testű, 9 mm-es állatok 4 pár tányérszerű tracheakopoltyúja a potroh hátoldalán helyezkedik el két sorban. A potrohszelvények hátoldalán gyakran páros fogszerű kiemelkedések figyelhetők meg. Testüket sokszor feltűnő világos és sötét vonalakból álló mintázat díszíti (23. és 31b-c. ábrák).

23. ábra (a-b) Serratella ignita (Ephemerellidae) lárvák – 9 mm

24. ábra (a) Erezett kérész (Ecdyonurus sp., Heptageniidae) lárva – 1 cm. (b) Epeorus assimilis (Heptageniidae) lárva – 1 cm

25. ábra (a-d) Hím dunavirág (Ephoron virgo, Polymitarcyidae) szubimágó vedlése imágóvá – 15 mm

26. ábra (a) Dánkérész (Ephemera danica, Ephemeridae) lárva – 3 cm. (b) Tiszavirág (Palingenia longicauda, Palingeniidae) lárvák – 3 cm. (c) Dunavirág (Ephoron virgo, Polymitarcyidae) lárva (15 mm) és feji része (kis kép)

27. kép (a) Folyamvirág (Potamanthus luteus, Potamanthidae) lárva – 17 mm. (b) Törpekérész (Caenidae) lárvák – 9 mm

28. ábra (a-b) Isonychia ignota (Isonychiidae) lárva – 2 cm. (c) Elevenszülő kérész (Cloeon dipterum, Baetidae) lárva – 6 mm

29. ábra (a) Elevenszülő kérész (Cloeon dipterum, Baetidae) lárvák – 6 mm. (b-c) Tavi kérész (Siphlonurus lacustris, Siphlonuridae) lárvák – 1 cm

30. ábra (a-b) Tavi kérész (Siphlonurus lacustris, Siphlonuridae) lárvák – 1 cm. (c) Metreletus balcanicus (Ameletidae) lárva – 11 mm

31. ábra (a) Paraleptophlebia sp. (Leptophlebiidae) lárva – 1 cm. (b) Serratella ignita (Ephemerellidae) lárva – 9 mm. (c) Torleya major (Ephemerellidae) lárva – 8 mm
Felhasznált és ajánlott irodalom
Andrikovics S, Fink TJ, Cser B (1992) Tiszavirág monográfia. Tisza Klub Füzetek, 2
Andrikovics S, Turcsányi I. (2001) Tiszavirág. Tisza Klub Füzetek, 10
Andrikovics S, Kerekes J, Kriska Gy, Liszi J (2002) A limnológia alapjai – főiskolai jegyzet. Eszterházy Károly Főiskola, Eger
Barber-James HM, Gattolliat JL, Sartori M, Hubbard MD (2008) Global diversity of mayflies (Ephemeroptera, Insecta) in freshwater. Hydrobiologia 595: 339–350
Barnes RD (1980) Invertebrate zoology, 4th edn. Saunders College, Philadelphia
Bauernfiend E (1994) Bestimmungsschlüssel für die Österreichischen Eintagsfliegen (Insecta: Ephemeroptera), 1. Teil. Wasser und Abwasser Suppl. 4/94:5-92
Bauernfeind E, Humpesch UH (2001) Die Eintagsfliegen Zentraleuropas (Insecta: Ephemeroptera): Bestimmung und Ökologie. Verlag des Naturhistorischen Museums, Wien
Bauernfeind E, Soldan T (2012) The mayflies of Europe (Ephemeroptera). Brill Academic Publishers, Leiden
Brittain JE (1982) Biology of mayflies. Annu. Rev. Entomol. 27: 119–147
Brittain JE (1990) Life history strategies in Ephemeroptera and Plecoptera. In: Campbell, IC (ed.) Mayflies and Stoneflies: Life Story and Biology. Kluwer Academic Publishers, Dordrecht, 1–12
Brittain JE, Sartori M (2009) Ephemeroptera. In: Resh VH, Cardé R (eds.) Encyclopedia of Insects, second ed. Academic Press, New York, 328–333
Buffagni A (1997) Mayfly community composition and the biological quality of streams. In: Landolt P, Sartori M (eds.) Ephemeroptera & Plecoptera Biology-Ecology-Systematics. Mauron+Tinguely & Lachat SA, Fribourg, 235–246
Buffagni A, Cazzola M, López-Rodríguez MJ, Alba- Tercedor J, Armanini DG (2009) Ephemeroptera. In: Schmidt-Kloiber A, Hering D (eds.) Distribution and ecological preferences of European freshwater organisms, vol. 3. Pensoft, Sofia-Moscow
Damborenea C, Rogers DC, Thorp JH (eds.) 2019 Keys to Palearctic Fauna: Thorp and Covich’s Freshwater Invertebrates. Vol. 4e4. Elsevier Inc.
Edmunds GF, McCafferty WP (1988) The mayfly subimago. Annu. Rev. Entomol. 33: 509–529
Egri Á, Pereszlényi Á, Farkas A, Horváth G, Penksza K, Kriska G (2017a) How can asphalt roads extend the range of in situ polarized light pollution? A complex ecological trap of Ephemera danica and a possible remedy. Journal of Insect Behavior 30: 374-384
Egri Á, Száz D, Farkas A, Pereszlényi Á, Horváth G, Kriska Gy (2017b) Method to improve the survival of night-swarming mayflies near bridges in areas of distracting light pollution. Royal Society Open Science 4: 171166
Elliott JM, Humpesch UH, Macan TT (1988) Larvae of the British Ephemeroptera. A key with ecological notes, vol 49. Freshwater Biological Association, Windermere
Farkas A, Száz D, Egri Á, Barta A, Mészáros Á, Hegedüs R, Horváth G, Kriska Gy (2016) Mayflies are least attracted to vertical polarization: A polarotactic reaction helping to avoid unsuitable habitats. Physiology and Behavior 163: 219-227
Fitter R, Manuel R (1986) Freshwater life. Collins, New York
Grant PM (2001) Mayflies as food. In: Dominguez E (ed.) Trends in Research in Ephemeroptera and Plecoptera. Kluwer Academic/Plenum Publishers, New York, 107–124
Horváth G, Kriska Gy (2008) Polarization vision in aquatic insects, polarization patterns of waters and polarized ecological traps. In: Lancaster J, Briers RA (eds) Aquatic insects: challenges to populations. CABI, Wallingford, 204–229
Horváth G, Kriska Gy, Malik P, Robertson B (2009) Polarized light pollution: a new kind of ecological photopollution. Front Ecol Environ 7(6): 317–325
Horváth G, Egri Á, Meyer-Rochow VB, Kriska Gy (2020) How did amber get its aquatic insects? Water-seeking polarotactic insects trapped by tree resin. Historical Biology 32: 1-11 (doi: 10.1080/08912963.2019.1663843)
Illies J (ed) (1978) Limnofauna Europaea. A checklist of the animals inhabiting European inland waters, with account of their distribution and ecology, 2nd revised and enlarged edn. G. Fischer/Swets & Zeitlinger, Stuttgart/Amsterdam
Jacobsen RE (1995) Symbiotic associations between Chironomidae (Diptera) and Ephemeroptera. In: Corkum LD, Ciborowski JJH (eds.) Current Directions in Research on Ephemeroptera. Canadian Scholars’ Press Inc., Toronto, 317–332
Kluge NJ (2004) The Phylogenetic System of Ephemeroptera. Kluwer Academic Publishers, Dordrecht
Kovács T, Ambrus A, Bánkuti K (1999) Data to the Hungarian mayfly (Ephemeroptera) fauna arising from collectings of larvae. Folia historico-naturalia Musei matraensis 23: 157–170
Kovács T, Péter J, István TP (2001) Ephemeroptera, Odonata and Plecoptera larvae from the River Tisza (1997–1999). Folia historico–naturalia Musei matraensis, 25: 135–143
Kovács T, Ambrus A (2001) Ephemeroptera, Odonata and Plecoptera larvae from the River Rába and Lapincs. Folia historico–naturalia Musei matraensis, 25: 145–162
Kovács T, Ambrus A, Bánkuti K, Juhász P (2003) Data to the Hungarian mayfly (Ephemeroptera) fauna arising from collectings of larvae II. Folia historico–naturalia Musei matraensis, 27: 59–72
Kovács T, Bauernfeind E (2003) Checklist of the Hungarian mayfly fauna (Ephemeroptera). Folia Entomol Hung 64: 69–84
Kriska Gy, Horváth G, Andrikovics S (1998) Why do mayflies lay their eggs en masse on dry asphalt roads? Water–imitating polarized light reflected from asphalt attracts Ephemeroptera. J Exp Biol 200: 2273–2286
Kriska Gy, Andrikovics S (1999) Phenological data on a parasitic relationship between Electrogena lateralis (Curtis, 1834) (Ephemeroptera) and Symbiocladius rhitrhogenae (Zavrel, 1924) (Chironomidae). Opuscula Zoological 31: 79–84
Kriska Gy (2004) Vízi gerinctelenek. Kossuth Kiadó
Kriska G, Bernáth B, Horváth G (2007) Positive polarotaxis in a mayfly that never leaves the water surface: polarotactic water detection in Palingenia longicauda (Ephemeroptera). Naturwissenschaften 94: 148-154
Kriska Gy. (2008) Édesvízi gerinctelen állatok – határozó. Nemzeti Tankönyvkiadó
Kriska Gy, Tittizer T (2009) Wirbellose Tiere in den Binnengewässern Zentraleuropas. Weissdorn, Jena
Kriska Gy (2013) Freshwater Invertebrates in Central Europe - A Field Guide. Springer–Verlag Wien Heidelberg New York Dordrecht London + Digital extra materials
Lampert K (1904) Az édesvizek élete. Királyi Magyar Természettudományi Társulat
Landa V, Soldán T, (1985) Phylogeny and Higher Classification of the Order Ephemeroptera: A Discussion from the Comparative Anatomical Point of View. CSAV, Praha
Málnás K, Polyák L, Prill É, Hegedüs R, Kriska Gy, Dévai Gy, Horváth G, Lengyel Sz (2011) Bridges as optical barriers and population disruptors for the mayfly Palingenia longicauda: An overlooked threat to freshwater biodiversity? Journal of Insect Conservation 15: 823-832
Mcmafferty WP (1998) Aquatic entomology. Jones and Bartlett, Bredbury
Mészáros Á, Kriska Gy. Egri Á (2021) Spectral optimization of beacon lights for the protection of night-swarming mayflies. Insect Conservation and Diversity, 14(2): 225-234
Mcgavin GC (ed) (2001) Insects. Oxford University Press, Oxford
Móczár L (1969) Állathatározó I. Tankönyvkiadó, Budapest
Munjiu O (2017) Distribution of endangered mayfly Palingenia longicauda (Olivier, 1791) (Ephemeroptera, Palingeniidae) on the territory of the Republic of Moldova. Lauterbornia, 84: 39-51
Nilsson AN (ed) (1996) Aquatic insects of North Europe. A taxonomic handbook, vol 1. Apollo Books, Stenstrup
Quigley M (1977) Invertebrates of stream and rivers, a key to identification. Edward Arnold, London
Sartori M, Brittain JE (2015) Order Ephemeroptera. In: Thorp J, Rogers DC (eds.) Ecology and General Biology: Thorp and Covich's Freshwater Invertebrates, Academic Press, 873–891
Schwab H (1999) Süßwassertiere. Klett, Stuttgart
Studemann D, Landolt P, Sartori M, Hefti D, Tomka I (1992) Ephemeroptera. Insecta Helvetica Fauna 9., Neuchâtel
Száz D, Horváth G, Barta A, Robertson BA, Farkas A, Egri Á, Tarjányi N, Rácz G, Kriska Gy (2015) Lamp-lit bridges as dual light-traps for the night-swarming mayfly, Ephoron virgo: Interaction of polarized and unpolarized light pollution. Public Library of Science ONE (PLoS ONE) 10 (3): e0121194
Thorp JH, Rogers DC (eds.) 2015 Ecology and General Biology: Thorp and Covich’s Freshwater Invertebrates. Vol. 1e4. Academic Press London
Ujhelyi S (1959) Kérészek (Ephemeroptera) – Magyarország Állatvilága. Akadémiai Kiadó, V/5
Varga Z, Rózsa L, Papp L, Peregovits L (szerk.) (2021) Zootaxonómia: Az állatvilág sokfélesége. Pars Kft., Nagykovácsi
Wichard W, Gröhn C, Seredszus F (2009) Aquatic insects in Baltic amber. Kessel: Remagen Verlag